
Главный закон роста численности
изолированной популяции
В основе любых моделей лежат некоторые предположения. Ценность модели определяется тем, насколько ее характеристики соответствуют свойствам моделируемого объекта. Одним из фундаментальных предположений, лежащим в основе всех моделей роста, является предположение о пропорциональности скорости роста численности популяции — самой этой численности, будь то популяция зайцев или популяция клеток. В основе этого предположения лежит тот общеизвестный факт, что важнейшей характеристикой живых систем является их способность к размножению.
Для многих одноклеточных организмов или клеток, входящих в состав
клеточных тканей — это просто деление, то есть удвоение числа клеток
через определенный интервал времени, называемый характерным временем
деления. Для сложно организованных растений и животных размножение
происходит по более сложному закону, но в наиболее простых и адекватных
моделях предполагается, что скорость размножения популяции
пропорциональна численности этой популяции.
Закон экспоненциального роста справедлив на определенной стадии роста для следующих живых систем: клеток в ткани, водорослей, бактерий в культуре, животных в популяциях. Математическое выражение, описывающее увеличение скорости изменения величины с ростом самой этой величины, называют автокаталитическим членом (авто — само, катализ — изменение скорости реакции).
Во многих популярных руководствах по экологии говорится, что экспоненциальный рост популяций возможен только в особо оптимальных условиях при отсутствии каких-либо ограничивающих факторов. Это не совсем верно, поскольку единственное необходимое и достаточное условие такого роста — это постоянство коэффициента естественного прироста, определяющего для размножающихся организмов скорость их размножения. Так, например, проводя серию наблюдений за ростом популяции каких-либо одноклеточных организмов в разных температурных условиях, нетрудно заметить, что с уменьшением температуры скорость деления клеток падает, но экспоненциальный характер роста сохраняется. [26]
Иногда желая принизить значение экспоненциального роста популяции, авторы акцентируют внимание на его непродолжительности, на то, что он почти никогда не встречается в природе и, следовательно, может рассматриваться, по их мнению, лишь как демонстрация потенциальных возможностей популяции к росту. При этом они забывают о том, что никакая популяция так бы никогда и не появилась в природе, если бы не существовал этот важный, пусть и кратковременный, этап ее развития. Но бывают случаи, когда этот этап все длится и длится и никак не может закончиться.
В 1859 г. один фермер завез в южную часть Австралийского континента дюжину кроликов из Европы. В Австралии для них не оказалось видов-контролеров (хищников или паразитов), и численность кроликов стала расти в соответствии с экспоненциальной кривой. В итоге за 6 лет количество кроликов достигло 22-х миллионов. К 1930 г. они расселились по всему континенту, а численность их достигла 750 млн.! Кролики конкурировали с овцами за корм (в итоге поголовье овец снизилось в два раза). Они лишали корма кенгуру. В начале 1950-х гг. удалось уничтожить 90 % кроликов, заразив их патогенным вирусом миксомы (родственником вируса оспы). Однако на этом «кроличья эпопея» в Австралии не завершилась: достаточно быстро произошел процесс формирования экотипа, устойчивого к болезни, и поголовье снова начало расти. [27]
В природе, прошедшей длительный путь эволюции, мы наблюдаем самые разнообразные способы ограничения экспоненциальной экспансии размножающихся организмов. Важное значение имеют внешние воздействия на популяцию: неблагоприятные условия, конкуренты, хищники, паразиты, возбудители болезней и т.д. Но для изолированных популяций интерес представляют только те изменения, которые возникают внутри самих популяций, происходящие в ответ на рост их численности.
Распространенное представление о том, что рост популяций в благоприятных условиях ограничивается только объемом пищевых ресурсов и конкуренцией — представляется ошибочным. Существует множество примеров, свидетельствующих о том, что все популяции: животные, растительные, бактериальные — обладают эффективными средствами, ограничивающими рост своей численности и активизирующимися задолго до того как заканчиваются пищевые ресурсы, или вступают в силу ограничения по причине конкуренции. Есть лишь редкие исключения из этого правила.
Такая саморегуляция, когда популяция ведет себя как единый живой организм, не является приобретением высших форм жизни. Она характерна для всех видов, даже бактериальных, вырабатывающих для этого целое семейство активных веществ. Высшие организмы регулируют свою численность множеством способов, например, через паразитов в составе биоценоза, пропуском сезонов размножения или даже рассасыванием беременности. [28]
Кажутся ли удивительными в таком случае парадоксальный гиперболический рост численности человеческой популяции и следующий за ним демографический переход, ограничивающий ее численность на некотором фиксированном уровне. Рост, который никогда не зависел ни от каких ресурсов и переход, который происходит в условиях всеобщего изобилия, когда нет (в первом приближении) никаких ограничений ни в пищевых, ни в пространственных, ни в энергетических, ни в каких-либо других ресурсах. Разве удивительно, что растущее человечество как система, с помощью разнообразных появляющихся и исчезающих связей, управляет своим ростом и ведет себя подобно всем другим видам и подобно Гее Лавлока, как единый живой организм?
То, что плотность популяции влияет на ее дальнейший рост можно проверить в опытах с любыми видами организмов. Например, при содержании белых мышей в вольерах, когда люди следят за чистотой клеток и обеспечивают всех кормом, мыши, достигнув определенной численности, перестают размножаться. Если перевести их в более просторную клетку, тем самым снизив плотность популяции, они продолжат размножение вновь до определенного предела. При этом меняются характер поведения мышей и отношения их между собой. Зверьки становятся беспокойными и агрессивными, и это отрицательно влияет на процесс размножения. [24]
Когда взаимодействие между особями изолированной популяции отсутствует, ее рост происходит по экспоненциальному закону. Этот закон был описан в книге Роберта Мальтуса «Опыт о законе народонаселения». В ней было впервые сформулировано положение о том, что численность популяции в благоприятных условиях растет по закону геометрической прогрессии. Сам русский термин «популяция» происходит от английского «population», население. Мальтус был первым, кто применил математику в экологии, если не считать итальянского математика Фибоначчи.
В своей работе Мальтус четко сформулировал необходимые идеализации, без которых была бы невозможна математическая постановка задачи: однородность и изолированность популяции, неограниченность ресурсов, постоянство коэффициентов рождаемости и смертности, отсутствие взаимодействия, способного нелинейно сказаться на приросте. Закон Мальтуса считается первым и самым важным законом экологии популяций. Законы экологии популяций, по мнению В.Л. Гинсбурга, напоминают законы физики [26].
«Закон Мальтуса описывает, как растут или
уменьшаются популяции, когда больше ничего не происходит. Он описывает
естественное состояние популяций – как они ведут себя в отсутствие
каких-либо внешних факторов (Гинзбург, Коливан 2004) ».
«Гинзбург (1986) заметил, что закон Мальтуса играет такую же роль в
экологии как Первый закон Ньютона в физике. До Галилея и Ньютона
Аристотель утверждал, что естественным состоянием тел является покой, а
движение возникает только тогда, когда к объекту приложена сила.
Господин Исаак Ньютон, однако, доказал, что верно обратное: постоянное
движение является естественным состоянием, а непостоянное движение и
покой возникают только тогда, когда к объекту приложена сила. Его первый
закон содержит концепцию инерции, которая является «стремлением тела
сопротивляться изменениям своей скорости (Кребс 2001б).»
«Подобно первому закону Ньютона, закон Мальтуса говорит о том, что
естественное состояние популяции – не покой (т.е. постоянная популяция),
а движение (т.е. экспоненциальный рост или уменьшение); и если популяции
не растут или уменьшаются экспоненциально, это происходит потому, что
внешняя сила (т.е. что-то в окружающей среде) изменяет уровень
рождаемости и / или смертности (Гинзбург 1986, Гинзбург, Коливан 2004).
Эта внешняя сила может быть как небиотическим так и биотическим фактором
как, например, «уровень межвидового заполнения и плотность всех
остальных видов в сообществе, которые могли бы взаимодействовать с
основными видами (Турчин 2003)».
Дадим определение закону экспоненциального роста сначала для колонии микроорганизмов, где смертность отсутствует, а затем и для произвольной популяции организмов:
Экспоненциальный, естественный (обусловленный только внутренними,
эндогенными, системными причинами, т.е. никак не управляемый не «извне»,
не «изнутри») рост численности популяции множества однородных
размножающихся организмов — это суперпозиция множества параллельных
процессов деления, размножения с постоянным коэффициентом естественного
прироста по закону одной и той же прогрессии на последовательности
временных интервалов одинаковой продолжительности, равной характерному
времени размножения, с равномерно распределенной фазой.
Размножающуюся популяцию можно представить как объединение элементарных, независимых, далее неделимых частиц, подсистем, состоящих, к примеру, из одной бактерии или пары разнополых представителей моногамной популяции. Т.е. эта частица, атом популяции, ее элементарная составляющая — «не видит», «не чувствует» других, размножается и гибнет независимо от них по закону геометрической прогрессии, одинаковому для всех.
В более сложном случае можно допустить взаимодействие такой элементарной подсистемы с другими, но лишь такое, которое оставляет неизменным коэффициент естественного прироста, вне зависимости от находящегося в системе числа «частиц».
Итак, главные условия экспоненциального роста численности популяции
это:
- Неизменность состояния среды (не обязательно строгая
неизменность, вариации возможны, но лишь в тех пределах, в
которых сохраняется гомеостаз организмов), в которой находится
популяция, следствием чего является строгая цикличность,
периодичность элементарного продуктивного процесса во времени.
Для экспоненциального роста колонии микроорганизмов, к примеру,
необходима неизменность концентрации питательной смеси, ее
температура, физические поля, в которых находятся организмы,
уровень радиации и т.д.
- Независимость, отсутствие взаимовлияния процессов размножения элементарных составляющих популяции, рассредоточенной в пределах среды обитания, результатом чего является аддитивность естественного прироста (скорости роста численности) любых ее подсистем. Колонию микробов, например, можно разбить на любые части, в которых будет разное число таких микробов, и скорость роста численности этой колонии будет равна сумме скоростей роста всех ее частей. Это свойство вытекает из линейности дифференциального уравнения (1).
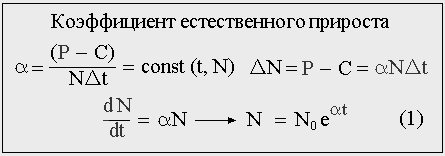
Рис 4. Главное условие экспоненциального роста популяции заключается в постоянстве коэффициента естественного прироста.
Главное условие экспоненциального роста заключается в том, что коэффициент естественного прироста популяции α, т.е. прирост ее численности за какой-то промежуток времени, отнесенный к этой численности, есть величина неизменная или «почти неизменная» в период роста (уменьшения) численности. Для популяции организмов со смертностью он равен разности между числом родившихся и числом умерших за единицу времени (Р-С), поделенную на общую численность. И число родившихся, и число умерших — случайные величины, различные по своей природе, имеющие разные математические ожидания и дисперсии и по разному меняющиеся во времени.
Коэффициент рождаемости (P/NΔt) и коэффициент смертности (C/NΔt) могут изменяться со временем в процессе роста популяции, но если при этом их разность будет оставаться неизменной — рост будет экспоненциальным. Если же это условие будет нарушено — экспоненты не получится; например, если для некоторой популяции коэффициент рождаемости — константа и не зависит от численности, а коэффициент смертности пропорционален численности, то рост будет логистическим.