
Частные случаи общего закона
Если в правой части уравнения (5) убрать и линейный и нелинейный член, то получим уравнение (7), в котором разность между числом родившихся и умерших за единицу времени (Р - С)/Δt равна нулю, и мальтузианский прирост, соответственно, будет равен убыли за счет взаимодействий. Нелинейный член может быть не монотонной функцией численности, и тогда рост будет более сложным. Устойчивый гомеостаз, когда численность популяции остается неизменной или слабо колеблется около положения равновесия, достигается при тех значениях численности, которые обращают правую часть уравнения (5) в нуль. И при которых вторая производная от правой части — отрицательна. [24]
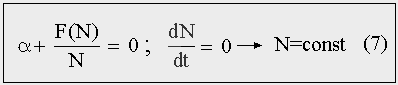
Рис. 7. Состояние популяционного гомеостаза.
Экспоненциальный рост возникает при отсутствии взаимодействий между членами популяции (он был рассмотрен нами ранее). В этом случае в обобщенном уравнении (5) необходимо отбросить нелинейный член F(N).
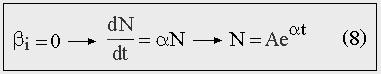
Рис. 8. Экспоненциальный рост.
Если для некоторой популяции коэффициент рождаемости есть величина постоянная и не зависит от численности, а коэффициент смертности пропорционален численности, то рост будет логистическим (9).
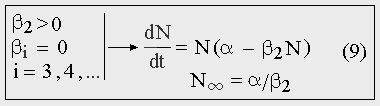
Рис. 9. Логистический рост.
Модель логистического роста, основанная на предположении об убывающей линейной зависимости удельной скорости популяционного роста от численности популяции, является наиболее простой из существующих моделей ограниченного роста. В этом ее несомненное достоинство. Но берется она, по сути, «с потолка», т.к. уравнение (9) никак не вытекает из каких-либо особенностей размножения и гибели организмов. Этим она отличается от модели естественного экспоненциального роста, которая полностью прозрачна и не содержит никаких искусственных предположений. Поэтому реальная S-образная кривая роста популяции может сильно отличаться от логистической кривой.
«Предположение о линейной зависимости скорости роста популяции от ее плотности (основное условие логистического роста) Ф. Смит (Smith, 1963) проверил экспериментально на лабораторной популяции рачка Daphnia magna. Увеличивая объем сосуда с питательной средой, в котором содержались дафнии, Ф. Смит в течение некоторого времени поддерживал плотность растущей популяции на одном уровне. Определив таким образом при разных плотностях значения удельной скорости популяционного роста, Ф. Смит построил по экспериментальным данным график, отражающий взаимосвязь данных величин. В соответствии с логистической моделью ожидалось, что график этот будет прямой линией, однако на самом деле получилась кривая, т. е. при низкой плотности популяция росла быстрее, чем это было бы при линейной зависимости, а при высокой — медленнее. Учтя эти данные и соответствующим образом модифицировав модель, Смит добился гораздо лучшего соответствия ее результатам эксперимента». [26]
В случае, когда нелинейный член (F(N) = -N2f(N)) представляет собой монотонную функцию можно говорить об обобщенном логистическом росте (9A). Такое уравнение называют иногда обобщенным уравнением роста Ричардса (Richards, Карманова, Иванилова, 1971) и используют для описания роста как отдельных организмов, так и популяций в целом. [26]
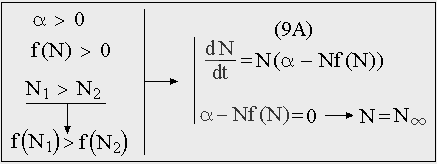
Рис. 10. Обобщенное уравнение логистического роста.
Какими чертами должны обладать организмы, чтобы рост их популяций описывался логистической или обобщенной логистической моделью? Во-первых, при возрастании численности (плотности) популяции должна возрастать вероятность их гибели и/или снижаться вероятность оставить потомство. Во-вторых, реакция этих организмов на возрастание численности (плотности), проявляющаяся в снижении рождаемости и/или увеличении смертности, должна осуществляться без запаздывания, иначе говоря, она должна быть значительно меньше времени жизни отдельной особи этой популяции, т.е. быть практически мгновенной.
Ни один реально существующий вид такими свойствами, очевидно, не обладает. И лучше всего такой модели соответствует рост простейших или бактерий, размножающихся в условиях конкуренции за пищевые ресурсы в среде, объем которой ограничен. Суть логистической модели заключается в том, что на начальной стадии роста, при малой численности, в правой части уравнения роста доминирует линейный член, и рост является экспоненциальным. По мере увеличения численности постепенно начинает сказываться присутствие квадратичного (нелинейного) члена, и численность устремляется устойчивым образом к некоторому предельному значению, величина которого зависит как от линейного, так и от нелинейного члена. Здесь важно то, что линейным членом, в отличие от нелинейного, в правой части логистического уравнения, пренебречь нельзя ни на какой стадии роста.
Если поменять знак второго члена логистического уравнения с минуса на плюс, получим некий антипод этого роста, когда связи между членами популяции способствуют, а не препятствуют приросту ее численности. Экспоненциальный рост, также справедливый здесь на начальном этапе, плавно переходит затем в гиперболический. (Еще раз подчеркнем, что речь здесь идет только о причинных законах роста!) Ни одна популяция в природе, в том числе и популяция Homo sapiens, как мы это сейчас покажем, по такому закону никогда не росла.
Более того, функция F(N) = -N2f(N), входящая в обобщенный причинный закон роста, для всякой свободно растущей в естественных природных условиях изолированной популяции должна быть, по-видимому, монотонно убывающей, принимающей лишь отрицательные значения функцией. Это так, поскольку связи между членами популяции, описываемые нелинейным членом F(N), определяют борьбу за территорию и/или ресурсы и всегда отрицательно сказываются на приросте ее численности. Такие связи не могут ускорить ее естественный экспоненциальный рост, а могут лишь его замедлить. Исключением из этого правила могла бы быть популяция Homo sapiens, единственная популяция в природе, представители которой обладают сознанием. На самом деле, как мы сейчас покажем, это не так, и рост населения Земли не может быть описан обобщенным причинным законом (5) с положительным нелинейным членом.
Действительно, «чисто гиперболический рост» населения мира, который, собственно, и был доказан в работах Форстера и Капицы, возможен лишь в том случае, если в разложении F(N) оставить только положительный квадратичный член β2N2, а линейным членом αN — пренебречь:
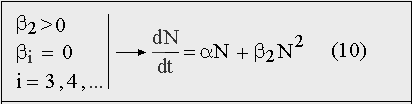
Рис. 11. Уравнение экспоненциального роста, переходящего в гиперболический, согласно которому не растет ни одна популяция в природе.
Но пренебречь членом αN для растущей популяции Homo sapiens нельзя из принципиальных соображений, поскольку человек всегда размножался так же, как и любой другой биологический вид, прежде всего, по закону Мальтуса: естественный прирост пропорционален численности. Нелинейный член β2N2, отвечающий за «прогрессивные взаимодействия» между членами человеческой популяции, мог лишь ускорить этот рост, но абсолютно доминировать при любой численности и на всех этапах роста, он не мог ни при каких обстоятельствах.
Иначе говоря, для того, чтобы рост был «чисто гиперболическим» на всех этапах этого роста, он должен был определяться на всех этих этапах только связями, взаимодействиями между членами популяции.
Причем не просто определяться; необходимо, чтобы связи описывались на всех этих этапах единственным и неизменным членом β2N2, стоящим в правой части уравнения роста с постоянным коэффициентом β2, не зависящим от растущей численности. При этом нужно учесть, что такое уравнение роста как причинный закон описывает простейшую нелинейную ПОС, т.е. неустойчивый итеративный процесс, приводящий тем не менее по каким-то непонятным причинам к устойчивому гиперболическому росту популяции.
В такое совершенно невозможно поверить, поскольку ничто не может отменить животной составляющей природы человека. Люди вовсе не роботы передающие, принимающие, перерабатывающие информацию и размножающиеся лишь благодаря коллективному взаимодействию Капицы. Составляющая прироста за счет рождаемости возникает по причине естественного человеческого желания иметь детей, глубоко «зашитого» в подсознании человека.
Если бы это было не так — не было бы и тех семи миллиардов людей, которые живут ныне на планете Земля. Действительно, и это трудно себе представить, что с тех пор как первый архантроп два миллиона лет тому назад перешел к прямохождению, стал пользоваться орудиями труда и проявил первые признаки разумности, сменилось 100000 (сто тысяч!) поколений. И на протяжении всего этого бесконечно долгого пути эволюции наши предки никогда «не забывали» о необходимости обзавестись потомством. (О жизнесберегающих технологиях, во все времена уменьшавших смертность и также влиявших на рост популяции, говорить здесь не будем.) И размножались они лишь благодаря какому-то коллективному взаимодействию? Можно ли в такое поверить!
Можно предвидеть следующее возражение: коллективное взаимодействие лишь сделало возможным этот чрезвычайно медленный рост численности наших предков, продолжавшийся сотни тысяч лет, рост, согласно которому не растут никакие популяции животных. И благодаря которому на момент начала неолита, т.е. спустя примерно 2 млн. лет после его начала, численность человечества возросла от ста тысяч до нескольких миллионов. Это была лишь некоторая добавка к мальтузианскому приросту, которой бы не было, не будь коллективного взаимодействия, а был бы обычный экспоненциальный, а затем логистический рост и гомеостаз с неизменной численностью в финале, определяемой существующими ООС, каким растут популяции животных того же класса, что и человек, и численность которых никогда не превышает примерно ста тысяч.
В том то и дело, что это не так. Утверждая, что гиперболический рост человечества начался 1,6 млн. лет тому назад, С.П. Капица фактически отбрасывает линейный член αN в уравнении роста (5), оставляя только нелинейную составляющую, отвечающую за взаимодействия. Следовательно, весь естественный прирост, а не какая-то прибавка к нему, происходит у него лишь благодаря влиянию (прямому или косвенному) коллективного взаимодействия. Но первые гоминиды мало чем отличались от человекообразных обезьян, и считать рост общей численности их популяций сначала линейным (-4,4 млн. лет, -1,6 млн. лет), а затем гиперболическим (-1,6 млн. лет, 1960 год) и происходящим лишь за счет связей, взаимодействий между членами популяций, как считает С.П. Капица, — представляется ошибочным.
«... Первые открытия принадлежат английскому антропологу Лики. В дальнейшем крупный вклад был сделан французской экспедицией, которой руководил Коппен, исследовавший раннюю эпоху становления человечества. Именно тогда начался гиперболический рост численности населения нашей планеты. (1,6 млн. лет тому назад) С тех пор эта численность увеличивалась прямо пропорционально квадрату населения мира вплоть до нашего времени, когда для гиперболического роста скорость обратно пропорциональна квадрату времени. Медленная в начале, по мере роста населения скорость все увеличивается и в итоге происходит быстрее, чем по экспоненте, устремляясь в бесконечность, в конечное время, равное Т1 = 2025 г.
Экспоненциальный рост предполагает только индивидуальную способность человека к размножению, которая не зависит от остальных людей. Поэтому в невзаимодействующей популяции экспоненциальный рост не зависит от суммарного населения, и в принципе рост определяется временем удвоения. Однако согласно новому пониманию роста человечества рост происходит в результате коллективного механизма умножения нашей численности. Причины этого могут быть разными, однако мы увидим, как коллективный механизм делает их эффективными факторами роста в масштабе всего человечества». Парадоксы роста
Какие из всего этого должны быть сделаны выводы? Т.к. мальтузианской
составляющей естественного прироста пренебречь нельзя ни на каком
этапе роста человеческой популяции, рост этот, очевидно, не мог быть
гиперболическим. Поскольку, как показывают многочисленные
исследования, — это все-таки не так, необходимо признать, что
дифференциальный закон, связывающий скорость роста и численность
населения Земли, не может считаться
причинным законом. И в соответствии с причинным законом dN/dt =
N2/C не растет ни одна популяция в природе.
Причины такого аномального гиперболического роста численности человечества заключены, по-видимому, в многочисленные возникающих и исчезающих управляющих связях, которыми был охвачен человеческий социум во все времена. И поскольку связи с такими парадоксальными свойствами, поднимающими естественный экспоненциальный рост численности населения Земли до уровня гиперболического, не могут возникать и поддерживаться сами по себе, то, очевидно, что они должны были обеспечиваться во все времена некой управляющей системой, стоящей над популяцией. И гиперболический рост населения Земли есть в таком случае рост «вынужденный», управляемый.