
Обобщенный закон роста численности
изолированной популяции
Каким должен быть закон свободного роста изолированной популяции в условиях среды, свойства которой не меняются и при учете взаимодействия ее членов? Будем считать выполненными все возможные идеализации рассмотренные нами ранее. Для такой популяции прирост за счет рождаемости, так же как и убыль за счет смертности, возрастает при увеличении ее общей численности. Поэтому в простейшем случае, без учета внутривидовых взаимодействий (тех из них, что влияют на прирост численности), скорость роста должна быть пропорционально общей численности. Поскольку даже и при учете взаимодействий, если их влияние устремить к нулю, обобщенный закон роста должен превращаться в уравнение Мальтуса, то дифференциальное уравнение этого закона должно быть уравнением первого порядка.
Процесс роста численности свободной популяции, т.е. популяции, рост которой никем и никак не регулируется, не зависит, при прочих равных условиях, от того на каком участке шкалы физического времени он наблюдается. Поэтому время, как независимая переменная, не должно явным образом входить в состав его правой части. Такие уравнения называются автономными. Структура правой части обобщенного закона должна иметь вид (5): линейный член αN плюс нелинейный член F(N), который описывает взаимодействие между членами популяции.
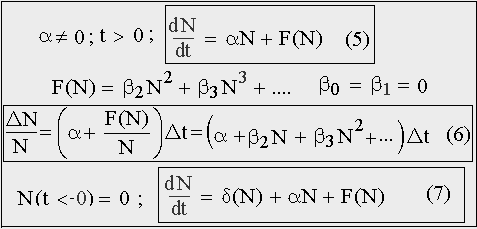
Рис. 5. Обобщенный закон свободного роста изолированной популяции.
Причем значение этой функции при N = 0 должно быть равным нулю: F(0) = 0, т.к. иначе пришлось бы допустить существование составляющей прироста, не зависящей от численности популяции. Так, например, при N = 0, т.е. при полном отсутствии членов популяции, скорость роста была бы не равна нулю. Что противоречит фундаментальному свойству жизни: живое происходит только от живого, и прирост определяется, прежде всего, численностью.
Если все таки допустить присутствие аддитивной константы в правой части уравнения роста, то в простейшем случае, если отбросить линейный и нелинейный член и оставить только константу, получим закон линейного роста численности от времени, который не может описывать рост никакой свободно растущей популяции, поскольку прирост здесь является постоянным и никак не зависит от растущей численности.
Что противоречит феноменологической теории С.П. Капицы, согласно которой скорость роста численности гоминид на первом этапе продолжительностью 2,8 млн. лет была постоянна и не зависела от их растущей численности.)
Если же оставить линейный член плюс константа от нелинейного, получим простейшее линейное неоднородное дифференциальное уравнение первого порядка с постоянными коэффициентами. В зависимости от знаков С и α имеется четыре варианта роста численности:

Рис. 6. Пример простейших линейных законов, которые не могут служить
для описания свободного роста (убывания) численности популяции.
Для случая С > 0, α < 0 можно, например, предложить такую легенду: имеется производство товаров с ограниченным сроком службы; С — число товаров производимых в месяц, αN — число товаров, вышедших из эксплуатации за этот месяц, — С/α — асимптота. Случай С > 0, α > 0 можно интерпретировать как свободный рост популяции с учетом постоянного дополнительного прироста за счет клонирования. Случай С < 0, α > 0 — рост численности популяции рыб в «неограниченном» водоеме с заданной квотой отлова. Последние два примера — это примеры несвободного, управляемого роста численности популяции. Итак, уравнение (4) не может служить для описания динамики свободного роста популяции каких-либо организмов из-за присутствия в его правой части аддитивной константы.
В дальнейшем будем говорить только о мальтузианской составляющей, определяющей рост популяции, т.е. считаем, что α > 0. Согласно теореме о разложении функции в степенной ряд, любую «достаточно хорошую» функцию всегда можно в такой ряд разложить. Поэтому нелинейный член F(N) в правой части уравнения (5) разлагаем в ряд Маклорена, при этом первый и второй член разложения должны быть равны нулю: βo = β1 = 0, т.к. константу отбрасываем, а линейный член равен αN, α > 0. Полученное уравнение с разделяющимися переменными можно проинтегрировать для каждой конкретной F(N). Отсутствие аддитивной константы в правой части приводит к тому, что она обращается в нуль при N = 0. Т.к. левая часть уравнения — это производная от численности по времени или скорость роста, то для кривой роста имеется горизонтальная асимптота, совпадающая с осью времени, т.е. такая же асимптота, как у экспоненты.
Это хороший показатель, он говорит о том, что рост численности популяции, определяемый обобщенным законом роста в его идеальном описании, с непрерывной численностью, не имеет начала. Если бы рост начинался в какой-то определенный момент времени, пришлось бы давать какое-то объяснение выделенности этого момента, как, например, при описании степенного параболического роста. Кроме того, очень важно понимать то, что линейным членом αN в обобщенном уравнении роста (5) пренебречь нельзя в принципе. Перечислим причины, почему это так:
- Т.к. разложение F(N) начинается с квадратичного члена, то F(N)/αN → 0 при N → 0, откуда следует, что при небольшой численности рост описывается линейным уравнением, является экспоненциальным и не зависит в первом приближении от взаимодействия между членами популяции. Т.е. получается правильная асимптотика.
- Если отбросить линейный член αN, оставить только F(N) и считать, например, что F(N) = βiNi, βj = 0, j ≠ i, т.е. все члены разложения кроме одного равны нулю, как в уравнении Капицы, то получаем причинный закон степенного роста, согласно которому, как мы показали ранее, не растет ни одна популяция. Если же в разложении F(N) присутствует более одного члена, а функция F(N) является монотонной, что соответствует любому реально возможному росту, то и в этом случае можно показать, что рост будет аналогичен степенному со всеми теми противоречиями, которые были рассмотрены нами ранее.
- Согласно первому закону экологии популяций, все популяции в неизменных и благоприятных внешних условиях и при отсутствии взаимодействий растут экспоненциально. Взаимодействия могут замедлить или ускорить этот экспоненциальный рост, но полностью отменить его они не могут. Если взять, к примеру, размножающееся человечество, то это, прежде всего, биологический вид, такой же, как и множество других видов, когда-либо существовавших в природе, умножающий свою численность по закону Мальтуса, и только затем его можно рассматривать как совокупность существ с множеством изученных и неизученных социальных связей, влияющих на все и вся, в том числе и на мировой естественный прирост.
Важным следствием обобщенного закона является зависимость (6): зависимость коэффициента естественного прироста (прироста численности, приходящегося на каждую особь популяции ΔN/NΔt за единицу времени) от полной численности этой популяции. Эта зависимость может иметь место только в том случае, если популяция представляет собой систему взаимодействующих особей, что возможно для сосредоточенной популяции с небольшим по площади ареалом обитания или для пространственно рассредоточенной, но объединенной единым информационным полем Мир-системы растущего человечества. Что полностью отвечает тем идеализациям, которые изначально закладывались в обобщенную модель. И что, несомненно, значительно снижает ее эвристическую ценность.
Учет «распространения» в пространстве особей (информации) приводит к необходимости построения моделей второго типа по нашей классификации, применения уравнений типа диффузия-кинетика, т.е. значительно более сложной математики.
Для рассредоточенной популяции животных и для человеческого общества это условие представляется слишком жестким и вряд ли может быть в реальности выполнено, т.к. размер ареала обитания популяции может в сотни раз превосходить расстояние, которое особь проходит за время своей жизни.
Кроме того, «сильно рассредоточенную» в пространстве популяцию вряд ли вообще можно считать популяцией по определению. Здесь может оказаться нарушенной главная из принятых идеализаций: одинаковые и неизменные внешние условия для всех ее частей. Разные части такой популяции могут в таком случае размножаться в разных природно-климатических условиях, иметь различные коэффициенты прироста и считаться отдельными популяциями.
А все человечество в целом вообще не представляло собой единое информационное поле ни в какие времена, исключая, возможно, последние два-три столетия. Вывод здесь такой: обобщенный причинный закон (5) по причине своей нелинейности имеет ограниченное применение и годится лишь для описания временно́й динамики изменения численности сосредоточенной, изолированной популяции. [25]
Наибольшую общность закону (5) можно придать, добавив в его правую часть дельта функцию Дирака (7), которая описывает акт творения или случайное зарождение жизни в первобытном океане Земли. Это «обобщение», впрочем, не нужно воспринимать слишком серьезно.